Mekanisme Pertahanan Tubuh
Sistem
kekebalan tubuh yang sehat merupakan kekebalan yang dapat membedakan antara
bagian tubuh dari sistem itu sendiri dan benda asing yang masuk ke dalam tubuh.
Secara garis besar, sistem imun menurut sel tubuh dibagi menjadi sistem imun
humoral dan sistem imun seluler. Sistem imun humoral terdiri atas antibodi dan
cairan yang disekresikan organ tubuh tubuh (saliva, air mata, serum, keringat,
asam lambung, pepsin, dan lain-lain). Sedangkan sistem imun dalam bentuk
seluler berupa makrofag, limfosit, dan neutrofil yang berada di dalam sel.

 Imunitas Alamiah terhadap Bakteri Ekstraselular
Imunitas Alamiah terhadap Bakteri Ekstraselular
Imunitas
Spesifik terhadap Bakteri Ekstraselular Netralisasi toksin
Netralisasi toksin
Sumber : https://duniahermanto.wordpress.com

 Imunitas Alamiah terhadap Bakteri Ekstraselular
Imunitas Alamiah terhadap Bakteri Ekstraselular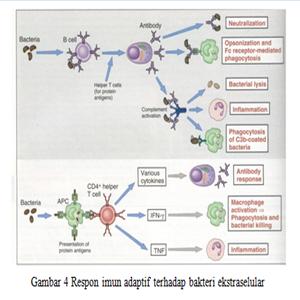 Netralisasi toksin
Netralisasi toksin
Komentar
Posting Komentar